The endocrine system is the system of glands, each of which secretes a type of hormone directly into the bloodstream to regulate the body.
The endocrine system is in contrast to the exocrine system, which secretes its chemicals using ducts. It derives from the Greek words "endo"
meaning inside, within, and "crinis" for secrete. The endocrine system is an information signal system like the nervous system, yet its effects and
mechanism are classifiably different. The endocrine system's effects are slow to initiate, and prolonged in their response, lasting for hours to weeks.
The nervous system sends information very quickly, and responses are generally short lived. Hormones are substances (chemical mediators)
released from endocrine tissue into the bloodstream where they travel to target tissue and generate a response. Hormones regulate various
human functions, including metabolism, growth and development, tissue function, and mood. The field of study dealing with the endocrine system
and its disorders is endocrinology, a branch of internal medicine.
Features of endocrine glands are, in general, their ductless nature, their vascularity, and usually the presence of intracellular vacuoles or granules storing their hormones. In contrast, exocrine glands, such as salivary glands, sweat glands, and glands within the gastrointestinal tract, tend to be much less vascular and have ducts or a hollow lumen.
In addition to the specialised endocrine organs mentioned above, many other organs that are part of other body systems, such as the kidney, liver, heart and gonads, have secondary endocrine functions. For example the kidney secretes endocrine hormones such as erythropoietin and renin.
The endocrine system is made of a series of glands that produce chemicals called hormones. A number of glands that signal each other in sequence is usually referred to as an axis, for example, the hypothalamic-pituitary-adrenal axis.
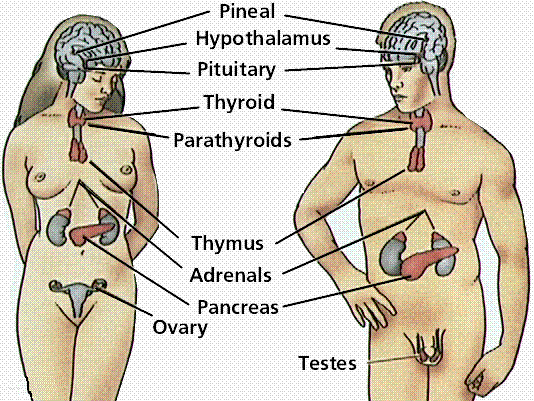
organs associated with endocrine system
control secretion rate click image to view animation The physiologic effects of hormones depend largely on their concentration in blood and extracellular fluid. Almost inevitably, disease results when hormone concentrations are either too high or too low, and precise control over circulating concentrations of hormones is therefore crucial.
The concentration of hormone as seen by target cells is determined by three factors:
Rate of production: Synthesis and secretion of hormones are the most highly regulated aspect of endocrine control. Such control is mediated by positive and negative feedback circuits, as described below in more detail.
Rate of delivery: An example of this effect is blood flow to a target organ or group of target cells - high blood flow delivers more hormone than low blood flow.
Rate of degradation and elimination: Hormones, like all biomolecules, have characteristic rates of decay, and are metabolized and excreted from the body through several routes. Shutting off secretion of a hormone that has a very short halflife causes circulating hormone concentration to plummet, but if a hormone's biological halflife is long, effective concentrations persist for some time after secretion ceases.
Feedback Control of Hormone Production
Feedback circuits are at the root of most control mechanisms in physiology, and are particularly prominent in the endocrine system. Instances of positive feedback certainly occur, but negative feedback is much more common.
Negative feedback is seen when the output of a pathway inhibits inputs to the pathway. The heating system in your home is a simple negative feedback circuit. When the furnace produces enough heat to elevate temperature above the set point of the thermostat, the thermostat is triggered and shuts off the furnace (heat is feeding back negatively on the source of heat). When temperature drops back below the set point, negative feedback is gone, and the furnace comes back on.
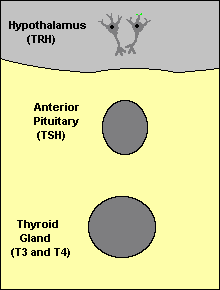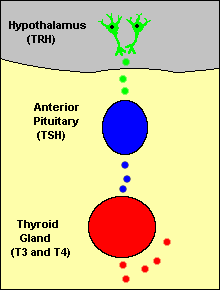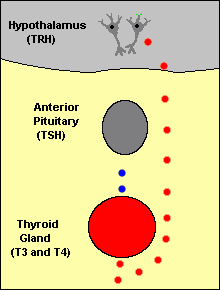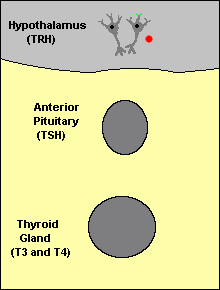
Feedback loops are used extensively to regulate secretion of hormones in the hypothalamic-pituitary axis. An important example of a negative feedback loop is seen in control of thyroid hormone secretion. The thyroid hormones thyroxine and triiodothyronine ("T4 and T3") are synthesized and secreted by thyroid glands and affect metabolism throughout the body. The basic mechanisms for control in this system (illustrated to the right) are:
Neurons in the hypothalamus secrete thyroid releasing hormone (TRH), which stimulates cells in the anterior pituitary to secrete thyroid-stimulating hormone (TSH).
TSH binds to receptors on epithelial cells in the thyroid gland, stimulating synthesis and secretion of thyroid hormones, which affect probably all cells in the body.
When blood concentrations of thyroid hormones increase above a certain threshold, TRH-secreting neurons in the hypothalamus are inhibited and stop secreting TRH. This is an example of "negative feedback".
Classes Of Receptors Membrane-bound receptors
these are proteins that are associated with the cell membrane. they can span across
the membrane and can transmit a signal from outside the cell to inside the cell.
outside the cell, a signal (like a hormone for example) will bind to the receptor.
This causes the receptor to change it's shape. The shape change is detected inside
the cell. It is the shape change that is the transmission of the signal from the outside
to the inside. Inside the cell, other proteins can interact with the receptor in it's new
shape and be turned 'on' to continue the signal pathway.
intracellular receptors
Intracellular receptors are receptors located inside the cell rather than
on its cell membrane. Examples are the class of nuclear receptors located
in the cell nucleus and the IP3 receptor located on the endoplasmic reticulum.
The ligands that bind to them are usually intracellular second messengers like
inositol trisphosphate (IP3) and extracellular lipophilic hormones like steroid
hormones. Some intracrine peptide hormones also have intracellular receptors.
Pituitary Gland
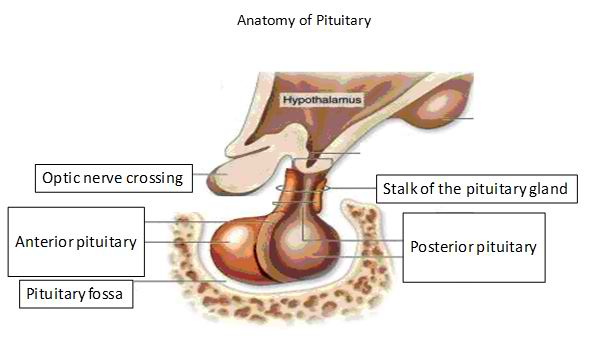
In vertebrate anatomy the pituitary gland, or hypophysis, is an endocrine gland about the size of a pea and weighing 0.5 g (0.02 oz.) in humans. It is a protrusion
off the bottom of the hypothalamus at the base of the brain, and rests in a small, bony cavity (sella turcica) covered by a dural fold (diaphragma sellae). The pituitary
is functionally connected to the hypothalamus by the median eminence via a small tube called the infundibular stem (Pituitary stalk). The pituitary fossa, in which the
pituitary gland sits, is situated in the sphenoid bone in the middle cranial fossa at the base of the brain. The pituitary gland secretes nine hormones that regulate homeostasis.
The pituitary gland consists of two components: the anterior pituitary (or adenohypophysis) and the posterior pituitary (or neurohypophysis), and is functionally linked to the hypothalamus by the pituitary stalk (also named the "infundibular stem", or simply the "infundibulum"). It is from the hypothalamus that hypothalamic tropic factors are released to descend down the pituitary stalk to the pituitary gland where they stimulate the release of pituitary hormones. While the pituitary gland is known as the 'master' endocrine gland, both of the lobes are under the control of the hypothalamus; the anterior pituitary receives its signals from the parvocellular neurons and the posterior pituitary receives its signals from magnocellular neurons.
Anterior pituitary (Adenohypophysis)
The anterior pituitary synthesizes and secretes the following important endocrine hormones:
Somatotropins:
Growth hormone (also referred to as 'Human Growth Hormone', 'HGH' or 'GH' or somatotropin), released under influence of hypothalamic Growth Hormone-Releasing Hormone (GHRH); inhibited by hypothalamic Somatostatin
Thyrotropins:
Thyroid-stimulating hormone (TSH), released under influence of hypothalamic Thyrotropin-Releasing Hormone (TRH)
Corticotropins:
Adrenocorticotropic hormone (ACTH), released under influence of hypothalamic Corticotropin-Releasing Hormone (CRH)
Beta-endorphin, released under influence of hypothalamic Corticotropin-Releasing Hormone (CRH)[3]
Lactotropins:
Prolactin (PRL), also known as 'Luteotropic' hormone (LTH), released under influence of multiple hypothalamic Prolactin-Releasing Factors (PRH) including dopamine, estrogen, progesterone and thyrotropin-releasing hormone.
Gonadotropins:
Luteinizing hormone (also referred to as 'Lutropin' or 'LH' or, in males, 'Interstitial Cell-Stimulating Hormone' (ICSH))
Follicle-stimulating hormone (FSH), both released under influence of Gonadotropin-Releasing Hormone (GnRH)
Melanotrophins
Melanocyte–stimulating hormones (MSHs) or "intermedins," as these are released by the pars intermedia, which is "the middle part"; adjacent to the posterior pituitary lobe, pars intermedia is a specific part developed from the anterior pituitary lobe.
These hormones are released from the anterior pituitary under the influence of the hypothalamus. Hypothalamic hormones are secreted to the anterior lobe by way of a special capillary system, called the hypothalamic-hypophysial portal system.
The anterior pituitary is divided into anatomical regions known as the pars tuberalis, pars intermedia, and pars distalis. It develops from a depression in the dorsal wall of the pharynx (stomodial part) known as Rathke's pouch.
Posterior pituitary (Neurohypophysis)
The posterior pituitary stores and secretes the following important endocrine hormones:
Magnocellular Neurons:
Oxytocin, most of which is released from the paraventricular nucleus in the hypothalamus
Antidiuretic hormone (ADH, also known as vasopressin and AVP, arginine vasopressin), the majority of which is released from the supraoptic nucleus in the hypothalamus
Oxytocin is one of the few hormones to create a positive feedback loop. For example, uterine contractions stimulate the release of oxytocin from the posterior pituitary, which, in turn, increases uterine contractions. This positive feedback loop continues throughout labor.
Thyroid Gland
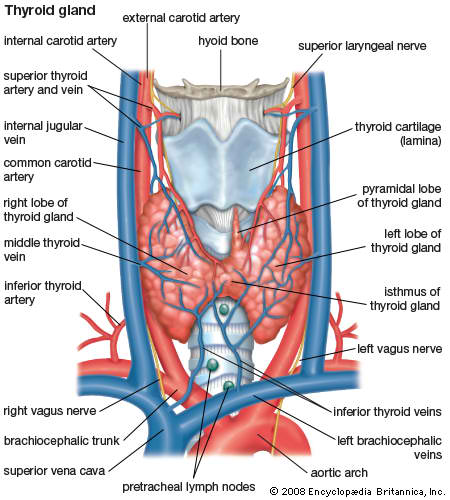
The thyroid gland or simply, the thyroid, in vertebrate anatomy, is one of the largest endocrine glands. The thyroid gland is found in the neck, below (inferior to) the thyroid cartilage (which forms the laryngeal prominence, or "Adam's apple"). The isthmus (the bridge between the two lobes of the thyroid) is located inferior to the cricoid cartilage.
The thyroid gland controls how quickly the body uses energy, makes proteins, and controls how sensitive the body is to other hormones. It participates in these processes by producing thyroid hormones, the principal ones being triiodothyronine (T3) and thyroxine which can sometimes be referred to as tetraiodothyronine (T4). These hormones regulate the rate of metabolism and affect the growth and rate of function of many other systems in the body. T3 and T4 are synthesized from both iodine and tyrosine. The thyroid also produces calcitonin, which plays a role in calcium homeostasis.
Hormonal output from the thyroid is regulated by thyroid-stimulating hormone (TSH) produced by the anterior pituitary, which itself is regulated by thyrotropin-releasing hormone (TRH) produced by the hypothalamus.
The thyroid gets its name from the Greek word for "shield", due to the shape of the related thyroid cartilage. The most common problems of the thyroid gland consist of an overactive thyroid gland, referred to as hyperthyroidism, and an underactive thyroid gland, referred to as hypothyroidism.
T3 and T4 regulation
The production of thyroxine and triiodothyronine is regulated by thyroid-stimulating hormone (TSH), released by the anterior pituitary. The thyroid and thyrotropes form a negative feedback loop: TSH production is suppressed when the T4 levels are high. The TSH production itself is modulated by thyrotropin-releasing hormone (TRH), which is produced by the hypothalamus and secreted at an increased rate in situations such as cold exposure (to stimulate thermogenesis). TSH production is blunted by somatostatin (SRIH), rising levels of glucocorticoids and sex hormones (estrogen and testosterone), and excessively high blood iodide concentration.
An additional hormone produced by the thyroid contributes to the regulation of blood calcium levels. Parafollicular cells produce calcitonin in response to hypercalcemia. Calcitonin stimulates movement of calcium into bone, in opposition to the effects of parathyroid hormone (PTH). However, calcitonin seems far less essential than PTH, as calcium metabolism remains clinically normal after removal of the thyroid (thyroidectomy), but not the parathyroids.
Adrenal Glands
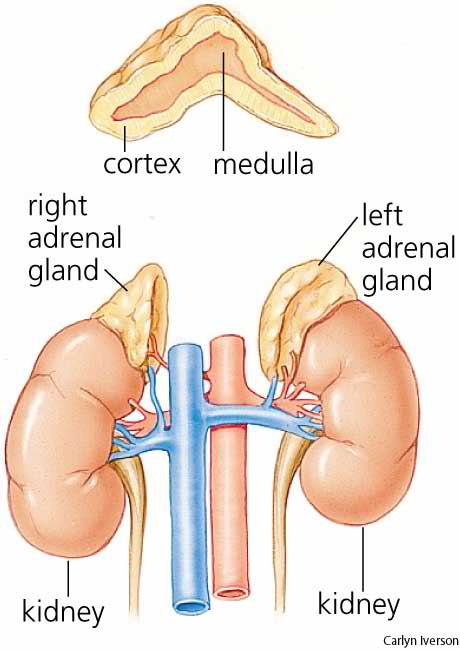
the adrenal glands (also known as suprarenal glands) are endocrine glands that sit atop the kidneys; in humans, the right suprarenal gland is triangular shaped, while the left suprarenal gland is semilunar shaped. They are chiefly responsible for releasing hormones in response to stress through the synthesis of corticosteroids such as cortisol and catecholamines such as epinephrine. The adrenal glands affect kidney function through the secretion of aldosterone, a hormone involved in regulating the osmolarity of blood plasma.
Anatomically, the adrenal glands are located in the retroperitoneum superior to the kidneys, bilaterally. They are surrounded by an adipose capsule and renal fascia. In humans, the adrenal glands are found at the level of the 12th thoracic vertebra. Each adrenal gland has two distinct structures, the outer adrenal cortex and the inner medulla, both of which produce hormones. The cortex mainly produces cortisol, aldosterone and androgens, while the medulla chiefly produces epinephrine and norepinephrine. The combined weight of the adrenal glands in an adult human ranges from 7 to 10 grams. Cortex
The adrenal cortex is devoted to the synthesis of corticosteroid and androgen hormones. Specific cortical cells produce particular hormones including aldosterone, cortisol, and androgens such as androstenedione. Under normal unstressed conditions, the human adrenal glands produce the equivalent of 35–40 mg of cortisone acetate per day. In contrast to the direct innervation of the medulla, the cortex is regulated by neuroendocrine hormones secreted from the pituitary gland which are under the control of the hypothalamus, as well as by the renin-angiotensin system.
The adrenal cortex comprises three zones, or layers. This anatomic zonation can be appreciated at the microscopic level, where each zone can be recognized and distinguished from one another based on structural and anatomic characteristics. The adrenal cortex exhibits functional zonation as well: by virtue of the characteristic enzymes present in each zone, the zones produce and secrete distinct hormones.
Zona glomerulosa (outer)
The outermost layer, the zona glomerulosa is the main site for production of mineralocorticoids, mainly aldosterone, which is largely responsible for the long-term regulation of blood pressure. Aldosterone's effects are on the distal convoluted tubule and collecting duct of the kidney where it causes increased reabsorption of sodium and increased excretion of both potassium (by principal cells) and hydrogen ions (by intercalated cells of the collecting duct). Sodium retention is also a response of the salivary ducts, distal colon, and sweat glands to aldosterone receptor stimulation. The major stimulus to produce aldosterone is angiotensin II while ACTH from the pituitary only produces a transient effect. Angiotensin is stimulated by the juxtaglomerular cells when renal blood pressure drops below 90 mmHg.
Zona fasciculata
Situated between the glomerulosa and reticularis, the zona fasciculata is responsible for producing glucocorticoids, such as 11-deoxycorticosterone, corticosterone, and cortisol in humans. Cortisol is the main glucocorticoid under normal conditions and its actions include mobilization of fats, proteins, and carbohydrates, but it does not increase under starvation conditions. Additionally, cortisol enhances the activity of other hormones including glucagon and catecholamines. The zona fasciculata secretes a basal level of cortisol but can also produce bursts of the hormone in response to adrenocorticotropic hormone (ACTH) from the anterior pituitary.
Zona reticularis
The inner most cortical layer, the zona reticularis produces androgens, mainly dehydroepiandrosterone (DHEA) DHEA sulfate (DHEA-S), and androstenedione (the precursor to testosterone) in humans.
Medulla
The adrenal medulla is the core of the adrenal gland, and is surrounded by the adrenal cortex. It secretes approximately 20% norepinephrine and 80% epinephrine. The chromaffin cells of the medulla, named for their characteristic brown staining with chromic acid salts, are the body's main source of the circulating catecholamines adrenaline (epinephrine) and noradrenaline (norepinephrine). Catecholamines are derived from the amino acid tyrosine and these water-soluble hormones are the major hormones underlying the fight-or-flight response.
To carry out its part of this response, the adrenal medulla receives input from the sympathetic nervous system through preganglionic fibers originating in the thoracic spinal cord from T5–T11. Because it is innervated by preganglionic nerve fibers, the adrenal medulla can be considered as a specialized sympathetic ganglion. Unlike other sympathetic ganglia, however, the adrenal medulla lacks distinct synapses and releases its secretions directly into the blood.
Cortisol also promotes epinephrine synthesis in the medulla. Produced in the cortex, cortisol reaches the adrenal medulla and at high levels, the hormone can promote the upregulation of phenylethanolamine N-methyltransferase (PNMT), thereby increasing epinephrine synthesis and secretion.
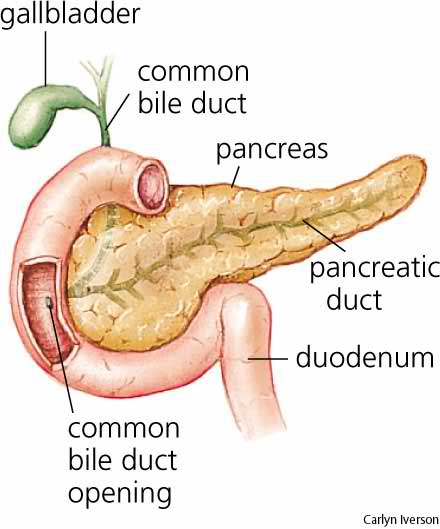
Pancreas
The pancreas is a gland organ in the digestive and endocrine system of vertebrates. It is both an endocrine gland producing several important hormones, including insulin, glucagon, and somatostatin, and a digestive organ, secreting pancreatic juice containing digestive enzymes that assist the absorption of nutrients and the digestion in the small intestine. These enzymes help to further break down the carbohydrates, proteins, and lipids in the chyme.
Under a microscope, stained sections of the pancreas reveal two different types of parenchymal tissue. Lightly staining clusters of cells are called islets of Langerhans, which produce hormones that underlie the endocrine functions of the pancreas. Darker-staining cells form acini connected to ducts. Acinar cells belong to the exocrine pancreas and secrete digestive enzymes into the gut via a system of ducts.
The pancreas is a dual-function gland, having features of both endocrine and exocrine glands.
The part of the pancreas with endocrine function is made up of approximately a million cell clusters called islets of Langerhans. Four main cell types exist in the islets. They are relatively difficult to distinguish using standard staining techniques, but they can be classified by their secretion: α cells secrete glucagon (increase glucose in blood), β cells secrete insulin (decrease glucose in blood), δ cells secrete somatostatin (regulates/stops α and β cells), and PP cells secrete pancreatic polypeptide.
The islets are a compact collection of endocrine cells arranged in clusters and cords and are crisscrossed by a dense network of capillaries. The capillaries of the islets are lined by layers of endocrine cells in direct contact with vessels, and most endocrine cells are in direct contact with blood vessels, either by cytoplasmic processes or by direct apposition. According to the volume The Body, by Alan E. Nourse, the islets are "busily manufacturing their hormone and generally disregarding the pancreatic cells all around them, as though they were located in some completely different part of the body." The islet of Langerhans plays an imperative role in glucose metabolism and regulation of blood glucose concentration.
The pancreas as an exocrine gland helps out the digestive system. It secretes pancreatic fluid that contains digestive enzymes that pass to the small intestine. These enzymes help to further break down the carbohydrates, proteins, and lipids (fats) in the chyme.
In humans, the secretory activity of the pancreas is regulated directly via the effect of hormones in the blood on the islets of Langerhans and indirectly through the effect of the autonomic nervous system on the blood flow.
Pineal Body
The pineal gland (also called the pineal body, epiphysis cerebri, epiphysis, conarium or the "third eye") is a small endocrine gland in the vertebrate brain. It produces the serotonin derivative melatonin, a hormone that affects the modulation of wake/sleep patterns and seasonal functions. Its shape resembles a tiny pine cone (hence its name), and it is located near the centre of the brain, between the two hemispheres, tucked in a groove where the two rounded thalamic bodies join. The pineal gland was originally believed to be a "vestigial remnant" of a larger organ. In 1917 it was known that extract of cow pineals lightened frog skin. Dermatology professor Aaron B. Lerner and colleagues at Yale University, hoping that a substance from the pineal might be useful in treating skin diseases, isolated and named the hormone melatonin in 1958. The substance did not prove to be helpful as intended, but its discovery helped solve several mysteries such as why removing the rat's pineal accelerated ovary growth, why keeping rats in constant light decreased the weight of their pineals, and why pinealectomy and constant light affect ovary growth to an equal extent; this knowledge gave a boost to the then new field of chronobiology.
Melatonin is N-acetyl-5-methoxy-tryptamine, a derivative of the amino acid tryptophan, which also has other functions in the central nervous system. The production of melatonin by the pineal gland is stimulated by darkness and inhibited by light. Photosensitive cells in the retina detect light and directly signal the SCN, entraining its rhythm to the 24-hour cycle in nature. Fibers project from the SCN to the paraventricular nuclei (PVN), which relay the circadian signals to the spinal cord and out via the sympathetic system to superior cervical ganglia (SCG), and from there into the pineal gland. The function(s) of melatonin in humans is not clear; it is commonly prescribed for the treatment of circadian rhythm sleep disorders.
The compound pinoline is also produced in the pineal gland; it is one of the beta-carbolines.
The human pineal gland grows in size until about 1–2 years of age, remaining stable thereafter, although its weight increases gradually from puberty onwards.The abundant melatonin levels in children are believed to inhibit sexual development, and pineal tumors have been linked with precocious puberty. When puberty arrives, melatonin production is reduced. Calcification of the pineal gland is typical in adults.
Apparently the internal secretions of the pineal gland inhibit the development of the reproductive glands, because in cases where it is severely damaged in children, the result is accelerated development of the sexual organs and the skeleton.In animals, the pineal gland appears to play a major role in sexual development, hibernation, metabolism, and seasonal breeding.